Migration introduces alleles two at once (BB diploid migrants vs A
Migration rates m >> Mutation rates u
|
|
WAA | WAB | WBB | qinit |
| Island | 1 | 1 - t | 1 - 2t | qi ~ 0 |
| Mainland | 0 | 0 | 1 | qm ~ 1 |
Migration rate m =
fraction of island
population newly arrived from mainland
[m equivalent to
fraction of new alleles arriving from mainland]
Migration / Selection equilibrium
resemble Mutation / Selection equilibrium
mathematically, except
Migration introduces alleles two at
once (BB diploid migrants vs A![]() B gametic mutations)
B gametic mutations)
Migration rates m >> Mutation
rates u
Calculate equilibrium frequency where ![]() qi
=
qi
= ![]() f(B) = 0 on island:
f(B) = 0 on island:
Change in f(B)
from migration: ![]() qi = (m)(qm
- qi)
qi = (m)(qm
- qi)
Change in f(B)
from selection: ![]() qi = -tqi(1
- qi)) / (1 - 2tqi)
qi = -tqi(1
- qi)) / (1 - 2tqi)
![]() -tqi(1 - qi)
[if tqi
<< 1]
-tqi(1 - qi)
[if tqi
<< 1]
Then,
combined change
![]() qi =
(m)(qm - qi)
- tqi(1 - qi)
qi =
(m)(qm - qi)
- tqi(1 - qi)
= mqm - mqi - tqi + tqi2)
= tqi2
- (m + t)(qi)
+ mqm
For ![]() qi = 0
solve as quadratic
equation for several special cases:
qi = 0
solve as quadratic
equation for several special cases:
if m ![]() t:
qi
t:
qi ![]()
![]() qm
migration
behaves
like mutation
qm
migration
behaves
like mutation
[except: alleles introduced at higher
rate m, as
diploids]
if m >> t:
qi ![]() qm
mainland B allele 'swamps' island A allele
qm
mainland B allele 'swamps' island A allele
if m
<< t:
qi ![]() (m / t)(qm)
some equilibrium
(m / t)(qm)
some equilibrium ![]() achieved, iff m constant
achieved, iff m constant
Intermediate cases can be simulated
in GSM worksheet
Approach to Mutation-Selection Equilibrium ![]() where
where
![]() qi
= 0
qi
= 0
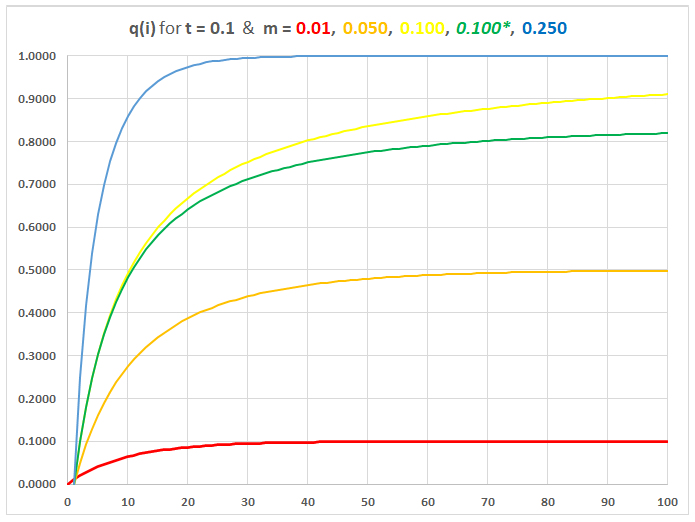
for t = 0.1 & m = 0.01
~ 0.250
For m = 0.250, qi rapidly approaches gm: migration from the Mainland swamps locally favored allele, prevents local adaptation.
For m = 0.01 or 0.05,
qi
reaches equilibrium at (m / t) as predicted:
deleterious mainland allele maintained at relatively high
frequency.
For m = t = 0.1, the
approximate solution m = 0.1
overestimates equilibrium qm.
The exact solution with m
= 0.1* is lower, but takes longer to achieve.