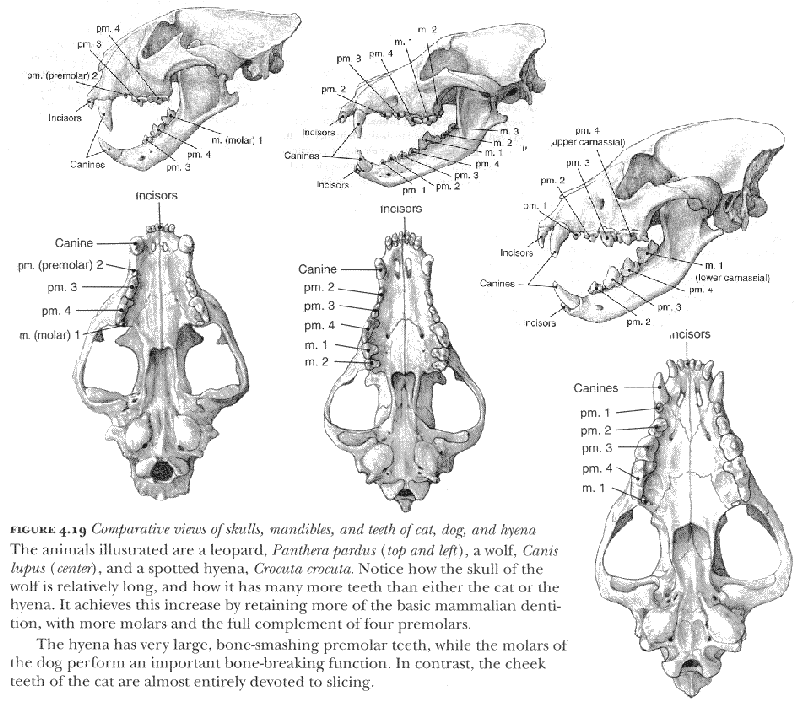
Since the cats are members of the Carnivora, their teeth exhibit many of the general features of the order, as may be seen in figure 4.19. But in comparison with, say, the dogs, all the cats have a very reduced dentition, that is, they have fewer teeth. That reduction comes about through specialization: whereas a dog is able to slice through meat and other relatively soft tissues and also retains the ability to crack bones, the cat is really only able to slice its food. Watching a domestic cat can be instructive here. Notice how it differs from a dog in the way it deals with its food, taking items from the bowl and chewing through pieces of meat on one side of its mouth, whereas the dog tends to put its muzzle in and bolt the lot down. You cannot feed a cat from the hand in the same way that you can a dog because each deals with its food in a different manner, and the cat usually drops the item to the floor before tackling it.
A dog will of course spend more time on eating if the food is hard or tough, or if it comes in larger pieces. Bones will be cracked and parts eaten, depending on the circumstances and the extent of hunger; wolves in the Canadian Arctic may return to a long~abandoned carcass in times of food shortage. Hyenas, especially the large spotted hyena, are even more adept at consuming bone and even better equipped to deal with it: their teeth permit them to break all but the largest items, and their digestive system can extract the entire organic fraction from the bone. In contrast, the cats generally show no real interest in crushing bones or extracting whatever nourishment is to be found in their marrow cavities, even though the larger cats are big and powerful enough to do considerable damage to bones. (Lions and tigers held in captivity may chew up much of the end of a large bone from a cow, but this activity may result as much from boredom as from any real desire to consume the bone.) It is however true that wild cheetahs have been observed to consume significant quantities of bones of very small prey, and to eat ribs and parts of the vertebrae of larger animals, so that capabilities and perhaps nutritional imperatives should not be overlooked.
If we examine the cheek teeth of a typical cat, whether it be a lion or a domestic tabby, we see that by far the most important feature is the scissorslike arrangement of the upper and lower carnassials, the meat-slicers. This arrangement is enhanced by the fact that the articulation for the mandible or lower jaw, the hinge, is in line with the intersection between the carnassials, just as in a pair of scissors. The other premolar teeth, although by no means unimportant to the animal, are relatively less significant. When we examine the cheek teeth of a saber-tooth such as Homotherium latidens, a species fairly common in Europe around 1.0 Ma ago, we see an even greater specialization in slicing, with the anterior check teeth much reduced in size.
The dogs have carnassials too, but they are only part of a dental armory that is augmented by many more premolars in front of the carnassials and by the crushing molars behind them. The dog is therefore a generalist when it comes to food-processing ability. In the hyenas the specialization is in almost entirely the opposite direction to the cats, with the development of huge, conical, bone-cracking teeth. Spotted hyenas in Africa today are capable of eating the entire carcass of a zebra, bones and all, but even they retain the carnassials to permit them to slice meat and other softer tissues.
Cats, then, are essentially flesh eaters: they eat primarily the meatier and softer parts of a carcass. But the mouth of a cat has a second notable feature in the size of its canine teeth. Those of the domestic cat are small but needle-sharp; those of a lion or tiger are massive conical structures---especially the upper ones, in which the unworn crown (the Dart that stands above the gum) may be up to 8 cm long in a large male. Such teeth are not merely for show, but are used extensively for subduing and killing prey once it is firmly held by the sharp claws; they may also be employed in fights between rivals. In older animals it is quite common to find one or more of the canines broken, or worn down to a smooth stump of variable length following a presumed break. Many such breaks are doubtless produced during prey capture, either by a struggling animal fighting to set itself free or, as in some cases that have been observed, when the cat is kicked in the face by a zebra, a large antelope, or even a giraffe). The canines also function, together with the incisors that lie between them in both upper and lower jaws, to help tear food off a carcass, and this action may lead to further wear and damage if bone is struck.
The saber-toothed cats carry the development of both the upper canines and the carnassials to extremes (but with important differences to be seen between the various lineages). However, it is as well to bear in mind, as we have already seen, that the saber-tooths were not unique, since at least three other groups among the mammalian meat eaters have developed their upper canines. Moreover, among our own order, the Primates, the upper canines of male baboons are fearsome-looking teeth, used extensively in display.
Among the saber-toothed felids the upper canines ... are usually elongated out from the jaw and flattened into bladelike shapes-hence the references to them in the literature as "saber"- or "dirk"- like. Such teeth often protrude below the lower jaw, or mandible, and in some cases are matched by a flange at the front of the mandible. The carnassials, both upper and lower, may become longer and narrower, like elongated scissors. In many cases the teeth between the canines and the carnassials are lost, become reduced in size, or develop their own bladelike characters, and in many species all three courses of evolution have been followed to some extent.
The cheek teeth of the saber-tooths therefore show a specialization toward slicing, one that is even further away from the limited bone-crushing abilities of the living cats. Such teeth must have been used with care if damage was to be avoided-and so too must the canines, in view of their length and slenderness. Numerous paleontologists over the years have therefore argued that subduing prey and killing it must have involved techniques that differed somewhat from those employed by the living cats. Many ideas have been proposed to explain the use of these enormously elongated teeth, with varying degrees of plausibility, but it is clear that they could not simply have been sunk deep into the neck of an animal in the process of leaping onto it and wrestling it to the ground, as many depictions have suggested: almost the slightest movement on the part of the struggling prey would have threatened breakage in such circumstances.
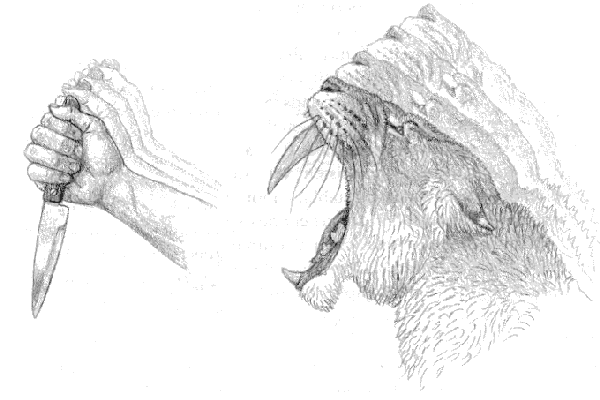
Strangely, the stabbing explanation in its various forms has tended to exclude the lower jaw (the mandible) from any mechanical calculations, simply carrying to extremes the analogy of the upper canines with a human wielding a steel knife (figure 4.22). Quite apart from the obvious risk of damage to the teeth, we see three main problems with this explanation. First, the machairodont canine is much more blunt than a steel knife. Second, driving such teeth deeply into the flesh of a victim in order to kill it during capture appears to us to require an excessive amount of force. Even if the necessary force could be generated through a combination of momentum and stabbing movements of the head, the risk of damage to the teeth through hitting bone or simple torsion during insertion would have been significant. Third, whatever the angle of attack chosen, it is difficult to see the mandible as other than an impediment to effective stabbing.
As an alternative to their use as a means of stabbing prey to death it has been suggested that the canines would have been mainly used to slice into the body of the prey in some manner, to make the flesh more accessible, once the animal had been killed. The first problem with that explanation is that it requires the prior assumption that the machairodonts existed largely as scavengers, since the method of killing prey is not considered.
Although virtually all living predators do scavenge to some extent, it is difficult to imagine that the machairodonts were generally able to adopt such a strategy in the face of competition for carcasses from the other large carnivores with which they coexisted. The second problem is that the wear on the canines, if they were habitually used in this way during feeding, would be greater than that generally observed: in many cases the canines are actually much less worn than the other teeth of individuals. It has also been pointed out that the sockets of the canines do not show the pattern of bony reinforcement about the opening that would be expected from such habitual use. As a living tissue, bone responds to stresses placed upon it, often by building more thickly where needed-in this case, where the bone emerges from the jaw. Instead, the sockets of the canines exhibit reinforcement toward the base, a pattern consistent with some kind of piercing action even if it fell short of a strongly stabbing motion.
What we seem to be left with as a general explanation is one put forward several years ago by the American paleontologist William Akersten, who suggested that a "shearing bite" to the abdomen of a temporarily immobilized prey might produce a gaping wound that would lead to significant blood loss and probably shock (figure 4.23). Akersten's model saw the enlarged flange at the front of the mandible as an anchor point against which the head depressor muscles might force the upper canines, producing a pierced rather than a stabbed wound. In the case of Smilodon in particular, he suggested that group action against juveniles of large species such as mammoths might even have been possible, with the group perhaps retreating for some time after the initial attack and waiting for the victim to die before returning to feed. Leyhausen describes how domestic cats may leave freshly dead prey for some time, and perhaps one could extend such behavior to machairodont kill techniques. But larger carnivores seem somewhat reluctant to lose contact with their prey once it is caught, and tend to retreat only if directly attacked, and we think it unlikely that a group of Smilodon would risk the seizure of their meal by other predators in the vicinity in such a way.
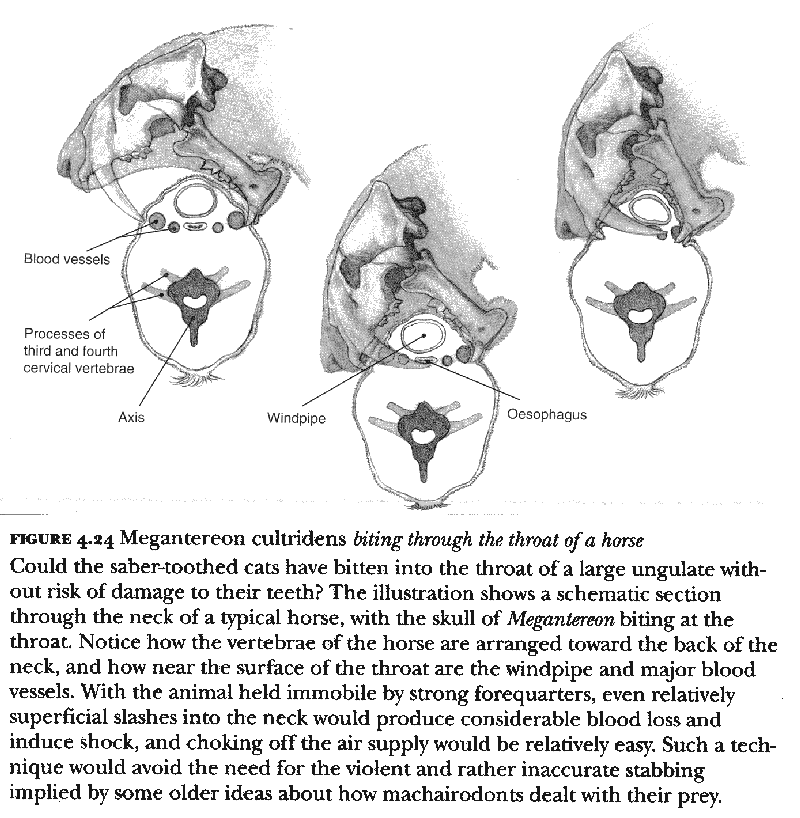
However, we do find Akersten's general model rather plausible, since it explains many features of the overall morphology of the machairodonts while avoiding the objections to other interpretations that rely in some way or other on the seemingly obvious analogy with an aggressive, stabbing mode of action. We also suggest that a shearing bite could be employed to the throat region of the neck of the prey, but only once the animal had been brought down and held virtually immobile, not while it was still standing (plate 9). Here the great strength that we see in the machairodonts, particularly in the forequarters of Megantereon, would have been important. As the illustrations in figure 4.24 show, once the prey is held still, it is possible to bite deeply into the throat, doing massive and, more importantly, rapid damage to the windpipe and major blood vessels.
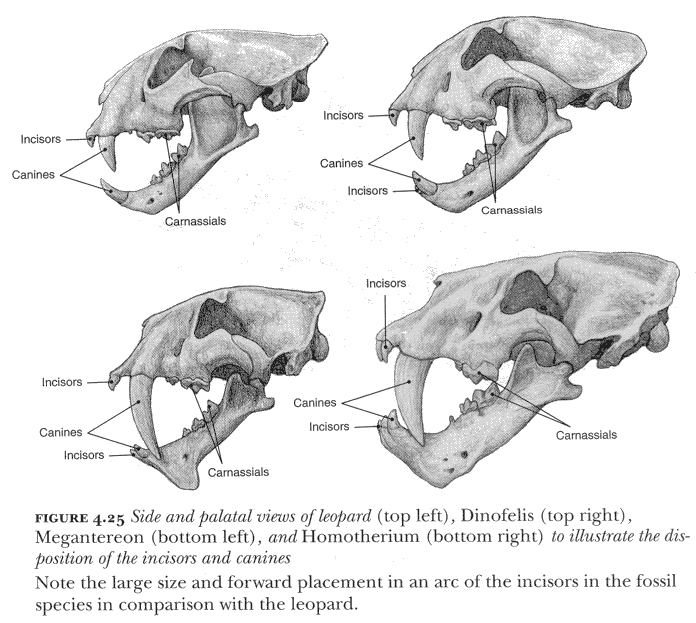
Whatever the true method of killing, the large size of the canines would also have meant that the method of getting food from a carcass differed from that employed by living cats, since they could not have been used in conjunction with the incisors to pull meat and skin away from the bones: they are simply too long. There is a further difference, however, exemplified in figure 4.25. In the living cats, such as the lion, tiger, and leopard, the six incisors in each upper and lower jaw are deployed as an essentially straight row of teeth between the left and right canines. The upper incisors are set somewhat forward of the line between the upper canines because the latter teeth actually sit somewhat behind and outside the lower canines when the mouth is closed, producing a gap into which the lower canine can fit. The central pair of incisors are the smallest, the two outside those are somewhat larger, and the two nearest the canines are largest of all. But even in a large lion the incisors are quite small teeth. In contrast, the incisors of many of the saber-toothed cats are often large, and the upper ones in particular are deployed in an arc that sets them well in front of the upper canines. This morphology is seen especially in the case of Homotherium, in which a view from the side shows the canine falling almost midway along the total extent of the dentition, and only slightly less so in Smilodon. In the case of Megantereon the protrusion of the upper incisor arc is also quite marked; and even in Dinofelis, with flattened but relatively short canines, the upper incisors are large. In other words, the incisor teeth in the sabertoothed species are developed and positioned in a way that overcomes the problem posed by the canines, and that permits the animal to get at and remove flesh from the bones.
In many ways the arrangements of the incisors in the saber-toothed cats are therefore more like those of a wolf or a hyena, as seen in figure 4. 19. Such a positioning might have allowed the animals to grasp the skin of their prey and to drag it backward, something that the large living cats may do-although this is perhaps not as efficient as moving a carcass by straddling it and walking forward with at least part of the body held off the ground.
{kind=link}
{kind=link}
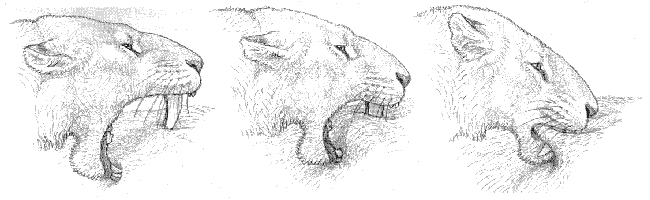{kind=link}
{kind=link}
{kind=link}